In my last post, I mentioned how highly homologous TRP channels respond in opposite ways to temperature changes. A clue to this paradox came later, when the structure of TRPA1 was published by the same lab as the TRPV1 structure(1). The previously discussed beta-bridges, or rather the segment that would form them if folded, is disordered in this structure. This is consistent with an "active", low-temperature state, which is also favored by disruption of the bridges--although as mentioned previously is controversial whether the specific human channel in this structure is cold-sensitive. In any case, given that highly homologous channels ARE unequivocally cold-activated, the necessary architecture should be present.
Lo and behold, the ARDs in the TRPA1 structure are oriented in a globally distinct fashion from in TRPV1, projecting perpendicularly to the membrane in a shape that I will call a "bullet" conformation--to distinguish it from the "pinwheel"--because of the combined shape of the ARDs plus the transmembrane domains.
Saturday, October 24, 2015
Tuesday, October 13, 2015
A TRP Down Structure Lane--Part 1
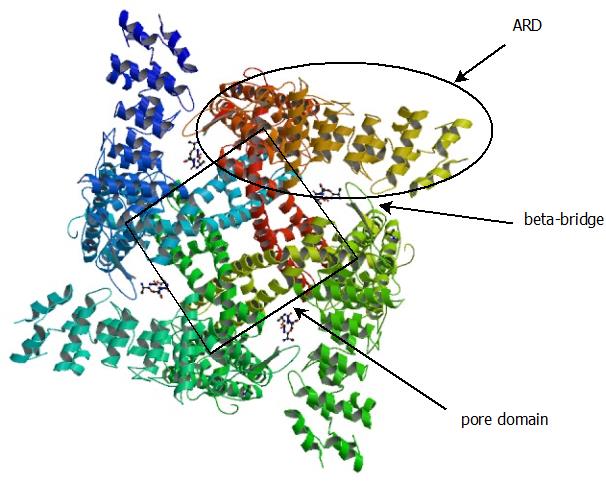
How do we know when a room is hot or cold? The family of transient receptor potential (TRP) ion channels senses general stimuli such as temperature, reactive chemicals, and pH, which open a cation-conducting pore. In particular, one member called TRPV1 is a very important part of the ability to sense painful heat, TRPM8 senses cool temperature, and TRPA1 senses electrophilic chemicals. TRPA1 also has a role as a painful cold sensor in some species, though the generality of this to humans is controversial(1), and in some species it is in fact a sensor of warmth(2). These channels are also regulated by more conventional ligands, for instance TRPV1 is activated by capsaicin from chili peppers (explaining why they "taste hot"), while TRPM8 is activated by menthol in mint.
How does a protein sense such general stimuli as temperature and chemical reactivity? In the case of electrophiles and pH, the answer is straightforward--these modify the chemical structure of the protein at nucleophilic and acidic sites, respectively, in a similar way as to how a chemical ligand binds to a receptor. In particular, cysteine and lysine side chains are the nucleophilic sites in TRPA1(3). Temperature is trickier--but the simplest mechanism is that there is a structural element in the protein that is ordered below a given temperature, and disordered above it. Other mechanisms involve physical or chemical changes to the membrane lipids--in particular, certain oxidized lipids were proposed to mediate heat sensation by TRPV1(4), although this is questionable(5).
The structure of TRPV1,
Tuesday, October 6, 2015
Back after a long hiatus
I decided to start this blog up again as a fully-specialized blog about science, leaning strongly toward discussions of structural biology. When I started this over 8 years ago, I was just out of undergrad and wanted a place to discuss all sorts of ideas, not just about science but about the world in general. And since my science-brain was developed but my worldview-brain wasn't (well, in many ways it still isn't, but that's another story), it was very disorganized. I'm sure most people who came here to read about science didn't care about my possibly weird opinions on everything that was going on in the news, society, etc.
(1)Kuhle B, Ficner R. A monovalent cation acts as structural and catalytic cofactor in translational GTPases. The EMBO journal. 33(21):2547-63. 2014. [pubmed]
Then why did I decide to start blogging again at all? Well, I'm finding that as I read through the many published articles on structural biology, I often have hypotheses and hunches that I could never publish, and that pertain to fields so far apart in terms of the underlying biology that I couldn't possibly study all of them. I felt I needed a place to "air" these ideas, where anyone who wants to read them (maybe even some people who work on the relevant systems) can do so.
While I sometimes have these ideas about other areas of science, I can't state them nearly as precisely as my hypotheses regarding protein structure. Therefore they're best suited to in-person discussions with me at a bar or something like that. Plus, being visual, I find that ideas about protein structure are more intuitive to describe, too.
An example of the why I want a blog like this--it turns out that the metal ion that I hypothesized here to trigger GTP hydrolysis in transcription factors has since been found to exist(1) (at least in eIF5, which is homologous to EF-Tu and EF-G), contrary to the preliminary data I reported on in my last post, and it's virtually exactly where I predicted it. It took an additional four years for it to be found, though, probably because the resolution of the ribosome-bound elongation factor structures was too low to observe it.
If you notice, I'm also moving to a more proper citation format. This post only has one, but upcoming posts may have many, and I want to make sure I'm properly referencing the publications on which I base my hypotheses.
Subscribe to:
Posts (Atom)