How do we know when a room is hot or cold? The family of transient receptor potential (TRP) ion channels senses general stimuli such as temperature, reactive chemicals, and pH, which open a cation-conducting pore. In particular, one member called TRPV1 is a very important part of the ability to sense painful heat, TRPM8 senses cool temperature, and TRPA1 senses electrophilic chemicals. TRPA1 also has a role as a painful cold sensor in some species, though the generality of this to humans is controversial(1), and in some species it is in fact a sensor of warmth(2). These channels are also regulated by more conventional ligands, for instance TRPV1 is activated by capsaicin from chili peppers (explaining why they "taste hot"), while TRPM8 is activated by menthol in mint.
How does a protein sense such general stimuli as temperature and chemical reactivity? In the case of electrophiles and pH, the answer is straightforward--these modify the chemical structure of the protein at nucleophilic and acidic sites, respectively, in a similar way as to how a chemical ligand binds to a receptor. In particular, cysteine and lysine side chains are the nucleophilic sites in TRPA1(3). Temperature is trickier--but the simplest mechanism is that there is a structural element in the protein that is ordered below a given temperature, and disordered above it. Other mechanisms involve physical or chemical changes to the membrane lipids--in particular, certain oxidized lipids were proposed to mediate heat sensation by TRPV1(4), although this is questionable(5).
The structure of TRPV1,
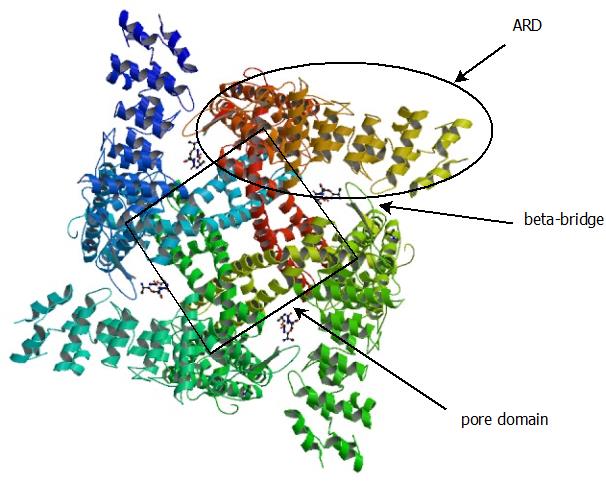
determined by cryoelectron microscopy, was published by the lab of David Julius at UCSF in December of 2013(6). It reveals the expected tetrameric architecture with a pore region and a cytoplasmic ankyrin repeat domain (ARD) on each subunit. The ankyrin repeat domains form a "four-bladed pinwheel" shape, with each ARD wrapping around toward its clockwise neighbor, as viewed from the cytoplasm (see picture above). Since cryo-EM is performed at temperatures far below the gating threshold of any TRP channel, cold-activated channels--or at least their temperature-sensing domains--would likely be in an active state, while heat-activated channels, including TRPV1, should be in an inactive state.
A quick glance at the structure reveals a likely element to unfold at high temperature. Near the junction between the ARD and the pore domain of each subunit, a beta-hairpin bridges the intersubunit gap to insert its tip into a pocket on the ARD of its neighbor. This hairpin, which I will call the "beta-bridge" from here on, makes few contacts to the rest of the protein, and is therefore likely prone to unfolding. Also, the character of the interaction makes sense for a sensor of noxious heat. The interaction primarily consists of interaction of a tyrosine and a proline on the hairpin with an aromatic cluster in the pocket, and noxious temperatures for biological systems correspond to those in which exposure of such hydrophobic groups to solvent occurs readily. As would be expected from this model, a segment including the beta-bridges was shown previously to be part of the heat sensor(7). This model also explains the regulation of TRPV1 by ATP, as the adenine base of ATP binds to a site on the ARD very close to the pocket into which the beta-bridge of the adjacent subunit docks(8).
These bridges constrain the ARDs in the orientation observed in the structure, but their disruption at high temperature would allow the domains to undergo rigid-body motion in any number of different directions. Such motion would be communicated to the pore, possibly through the C-terminal helices of the pore domain, which are encircled by the ARDs, and whose extensions form beta-strands that run along the beta-bridges. Which direction they move is anyone's guess, but given the symmetry of the structure and the way the ARDs wrap in a pinwheel shape, one likely possibility seems to be for them to swing outward in an "iris" type motion, becoming more radially oriented (see image below). This resembles the "square-to-windmill" transition observed for the inositol triphosphate receptor in response to calcium(9), with the caveat that this was not proposed as the motion leading to opening of that channel.

Now, that's all well and good(10), but what about cold-activated TRP channels like TRPM8 and TRPA1? If they worked the same way, the unfolding of their beta-bridges would need to be coupled to the closing of the channel. This would either require that they be oriented in a completely different manner, to create a linkage that "pulls" the ARDs the opposite way, or the mechanical "gearing" between the ARDs and the pore would have to be reversed. Either would be possible in principle, but it so happens that some of the electrophile-modified cysteines in TRPA1--which are much more conserved than the response to cold--lie in the region homologous to the beta-bridges. Given that electrophilic adducts on these cysteines may be any size and shape, it only makes sense for them to break an interaction, not form one. So, this would suggest that the coupling of beta-bridge detachment to activation is conserved from TRPV1 to TRPA1, regardless of the direction of the temperature response. More about this will be discussed in the next blog post.
(1)Chen J, Kang D, Xu J. Species differences and molecular determinant of TRPA1 cold sensitivity. Nature communications. 4:2501. 2013. [pubmed]
(2)Zhong L, Bellemer A, Yan H. Thermosensory and nonthermosensory isoforms of Drosophila melanogaster TRPA1 reveal heat-sensor domains of a thermoTRP Channel. Cell reports. 1(1):43-55. 2012. [pubmed]
(3)Hinman A, Chuang HH, Bautista DM, Julius D. TRP channel activation by reversible covalent modification. PNAS. 103(51):19564-8. 2006. [pubmed]
(4)Patwardhan AM, Akopian AN, Ruparel NB. Heat generates oxidized linoleic acid metabolites that activate TRPV1 and produce pain in rodents. The Journal of clinical investigation. 120(5):1617-26. 2010. [pubmed]
(5)De Petrocellis L, Schiano Moriello A, Imperatore R, Cristino L, Starowicz K, Di Marzo V. A re-evaluation of 9-HODE activity at TRPV1 channels in comparison with anandamide: enantioselectivity and effects at other TRP channels and in sensory neurons. British journal of pharmacology. 167(8):1643-51. 2012. [pubmed]
(6)Liao M, Cao E, Julius D, Cheng Y. Structure of the TRPV1 ion channel determined by electron cryo-microscopy. Nature. 504(7478):107-12. 2013. [pubmed]
(7)Yao J, Liu B, Qin F. Modular thermal sensors in temperature-gated transient receptor potential (TRP) channels. Proceedings of the National Academy of Sciences of the United States of America. 108(27):11109-14. 2011. [pubmed]
(8)Lishko PV, Procko E, Jin X, Phelps CB, Gaudet R. The ankyrin repeats of TRPV1 bind multiple ligands and modulate channel sensitivity. Neuron. 54(6):905-18. 2007. [pubmed]
(9)Hamada K, Terauchi A, Mikoshiba K. Three-dimensional rearrangements within inositol 1,4,5-trisphosphate receptor by calcium. The Journal of biological chemistry. 278(52):52881-9. 2003. [pubmed]
(10)I should point out that three structures of TRPV1 were published, one inactive, and two with agonists, capsaicin and resiniferatoxin. The pore shows conformational changes, and thus presumably the channels were responsive to the agonists even at cryogenic temperatures. However, the beta-bridges are still folded in these "active" structures, and though the ARDs move slightly between the structures, there is no clear displacement coordinate that distinguishes the two open states from the closed one. It appears that the coupling between the ARDs and the pore may be rather loose, and that chemical agonists may act by a local effect on the pore, rather than a global rearrangement of the subunits.
No comments:
Post a Comment